 English
English Español
Español
[Fungi on Mars? Evidence of Growth and Behavior From Sequential Images, published in Advances in Microbiology, vol 11, # 5. May 2021 original article here].
11 authors, including:

Rhawn Gabriel Joseph
Cosmology.com
88 PUBLICATIONS 1,873 QUOTES
View profile

Xinli Wei
Institute of Microbiology Chinese Academy of Sciences
64 PUBLICACIONES 322 CITAS
Ver Perfil

Ashraf M. T. Elewa
Minia University
83 PUBLICACIONES 628 CITAS
Ver Perfil
Olivier Planchon
French National Centre for Scientific Research
341 PUBLICACIONES 733 CITAS
Ver Perfil
Some of the authors of this publication are also working on these related projects: Life on Mars
Fungi on Mars? Evidence of Growth and Behavior From Sequential Images
R.G.Joseph.*1, Richard Armstrong2, Xinli Wei3, Carl Gibson4, Olivier Planchon5, David Duvall1,6, Ashraf M. T. Elewa7, N. S. Duxbury8, H. Rabb9, Khalid Latif10, Rudolph Schild*1,11
- Cosmology.com
- School of Life & Health Sciences, Aston University, Birmingham, UK.
- State Key Laboratory of Mycology, Institute of Microbiology, Chinese Academy of Sciences. Beijing. China
- Scripps Center for Astrophysics Space Sciences, Dept. Aerospace Engineering, University of California, San Diego, CA
- National Center for Scientific Research, Biogeosciences, University of Bourgogne, France
- Dept. of Zoology, Oklahoma State University, Emeritus, OK
- Geology Department, Faculty of Science, Minia University, Egypt
- Dept. of Physics and Astronomy, George Mason University, VA
- Grupo de Investigación en Astrobiología, Finlandia
- Centro Nacional de Excelencia en Geología, Universidad de Peshawar, Khyber Pakhtunkhwa, Pakistán
- Centro de Astrofísica, Harvard-Smithsonian, Cambridge, MA
Extract
Fungi thrive in radiation intense environments. Sequential photos document that fungus-like Martian specimens emerge from the soil and increase in size, including those resembling puffballs (Basidiomycota). After obliteration of spherical specimens by the rover wheels, new sphericals--some with stalks--appeared atop the crests of old tracks. Sequences document that thousands of black arctic “araneiforms” grow up to 300 meters in the Spring and disappear by Winter; a pattern repeated each Spring and which may represent massive colonies of black fungi, mould, lichens, algae, methanogens and sulfur reducing species. Black fungi-bacteria-like specimens also appeared atop the rovers. In a series of photographs over three days (Sols) white amorphous specimens within a crevice changed shape and location then disappeared. White protoplasmic-mycelium-like-tendrils with fruiting-body-like appendages form networks upon and above the surface; or increase in mass as documented by sequential photographs. Hundreds of dimpled donut-shaped “mushroom-like” formations approximately 1mm in size are adjacent or attached to these mycelium-like complexes. Additional sequences document that white amorphous masses beneath rock-shelters increase in mass, number, or disappear and that similar white-fungus-like specimens appeared inside an open rover compartment. Comparative statistical analysis of a sample of 9 spherical specimens believed to be fungal “puffballs” photographed on Sol 1145 and 12 specimens that emerged from beneath the soil on Sol 1148 confirmed the nine grew significantly closer together as their diameters expanded and some showed evidence of movement. Cluster analysis and a paired sample ‘t’ test indicates a statistically significant size increase in the average size ratio over all comparisons between and within groups (P = 0.011). Statistical comparisons indicates that arctic “araneiforms” significantly increased in length in parallel following an initial growth spurt. Although similarities in morphology are not proof of life, growth, movement, and changes in shape and location constitute behavior and support the hypothesis there is life on Mars.
Key Words: Fungi, Moulds, Puffballs, Geysers, Mud Volcanoes, Martian Mushrooms, Araneiforms, Mars, Life,
No Conflicts of Interest / Ethical Statement: All authors contributed and report no conflict of interest. *Corresponding authors: RhawnJoseph@gmail.com, rschild@cfa.harvard.edu
1. Introduction
Throughout its mission in Meridini planum, Mars, the rover Opportunity photographed chalky-white colored spherical shaped specimens upon the surface (Figures 7-16). Although unable to directly examine these spherical specimens, even misidentifying a flat oblong rock as a sphere (Bell et al. 2004), the Opportunity team published a number of articles claiming they are hematite (Christensen et al. 2004; Klingelhöfer et al. 2004; Soderblom et al. 2004; Squires et al. 2004). The hematite interpretation was immediately challenged and dismissed by Burt, Kaught and colleagues (Burt et al. 2005; Knauth et al. 2005) because of the “many contradictions” and pointing out that “unlike all known terrestrial concretions... they are uniformly spherical... uniform in their size distribution...” and concluding that: “The frequent analogy to hematitic spheroids is inappropriate" (see also Knauth et al. 2005). Two members of the Opportunity team also later admitted that much of the data was a “poor fit” for hematite (Glotch and Banfield 2006) whereas Bell et al (2004) acknowledged the data was "not consistent" with solid hematite but jarosite, and the spectral signatures obtained were indicative of “crystalline ferric iron.” The spherical hematite explanation was also rejected in 2006, by Joseph who instead coined the term “Martian mushrooms” while pointing out that these spherical specimens resemble lichens and mushrooms. In 2008 and 2014 Joseph again speculated that these spherical concretions may have been produced secondary to biological activity. Other scientists have also embraced a biological explanation for these spheroids (Dass, 2017; Kazmierczak 2020; Rabb 2015, 2018; Small 2015). In one study, dozens of experts in fungi and lichens identified these spheres as “puffballs”—a white spherical fungus belonging to the phylum Basidiomycota (Dass, 2017; Joseph 2016). Moreover, the rover Opportunity, in one three day sequence, photographed 9 spherical specimens that increased in size and an additional 12 that emerged from beneath the soil (Sols 1145 and 1148). The possibility that wind may have uncovered these specimens has been analyzed, discussed, and ruled out (Joseph et al 2020a,b). Many of these ground-level spherical specimens also have stalks or have shed portions of their outer membranes--possibly crustose--and are surrounded by white chunks and fluffy spore-like material that may consist of leprose (Figures 9, 11, 15, 16). In addition, amorphous white fungi-like specimens have been photographed in Gale Crater by the rover Curiosity within rock shelters. As exemplified by sequential images, these amorphous specimens expand in size, or conversely, change shape, move to new locations, and/or wane in size and nearly disappear (Figures 7, 17-21). In the same vicinity, and on the same days, mm-in-size dimpled-donut-shaped specimens that resemble mushrooms and lichen genus Ochrolechia, have also been photographed, some of which are attached to networks of white mycelium-like protoplasmic tendrils that snake along or above the surface (Figures 22-27). Some of these tendrils also increase in mass as documented by subsequent photos (Figure 22). However, if these “tendrils” are living, calcified, fossilized, or completely abiogenic is unknown. Massive black “araneiforms” (Figures 1-6, 45-51), hypothesized to consist of huge colonies of mould, fungi, algae, lichens, and bacteria (Ganti et al. 2003; Joseph et al. 2020c; Kereszturi et al. 2012; Ness, 2001) have also been photographed from orbit, appearing in the Spring, increasing in size, sometimes attaining a width and length up to several hundred meters only to wane and disappear with the coming of Autumn and Winter; and then the pattern is repeated the following Spring (Hansen et al. 2010; Kieffer et al. 2006; Lovett 2000; Portyankina et al. 2020; Prieto -Ballesteros, et al. 2006). Furthermore, photo sequences show the “growth” of what appear to be mould, bacteria, and fungi on the rovers Curiosity and Opportunity (Figures 36-39). Thus, in the Equatorial and Arctic regions, specimens and formations emerge, multiply, wax and wane in size and move or expand into new locations; actions that are characteristic of living organisms.
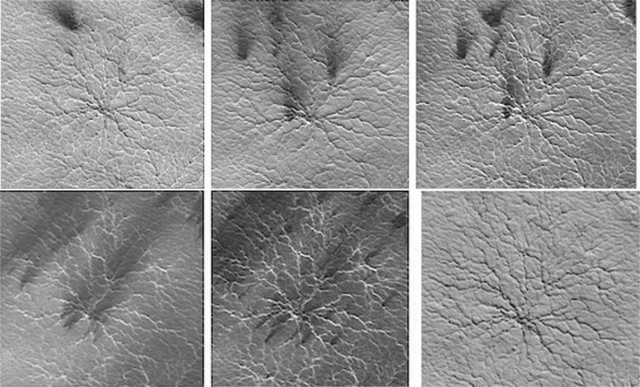
Figure 1. Mars orbital photos. Clockwise from Top: Early Spring. Mid Spring. Late Spring. Early Summer. Mid Summer. Autumn.
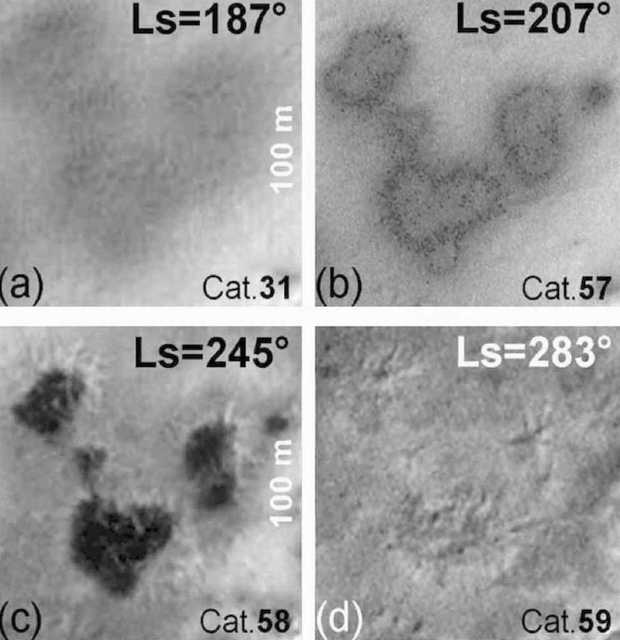
Figure 2. Waxing from gray in the (a) Winter to black during the (b,c) Spring and waning and disappearing in (d) Autumn/Winter. Photographed via satellite, from orbit (reproduced from Ganti et al. 2003).
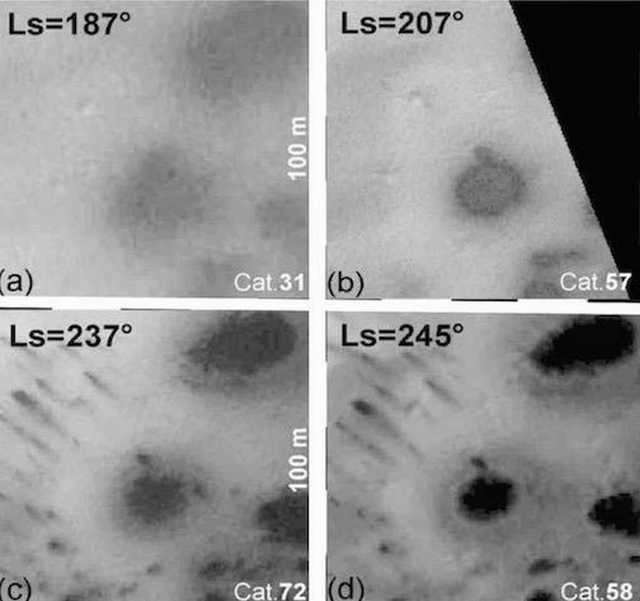
Figure 3. Waxing from gray in the Winter (a) to black during the Spring and early Summer (b-d). Photographed via satellite, from orbit (reproduced from Ganti et al. 2003).

Figure 4. Mars HiRISE orbital photos. (Top Left) Day 1. (Bottom) Day 34. Linear specimens travel up and over terrain with those in the center growing toward each other. Final length of dark linear specimens is 130 meters, with an estimated growth rate of 5 to 10 meters per day.

Figure 5. Mars HiRISE orbital photos. (Top Left) Day 1. (Central) Day 22. (Right) Day 34. Blotchy and linear specimens emerge from dark blotches at the top of this Martian dune. The linear specimens appear to grow outward and downward. Final length of dark linear specimens is estimated at 60 meters, with an estimated growth rate of 5 to 10 meters per day.
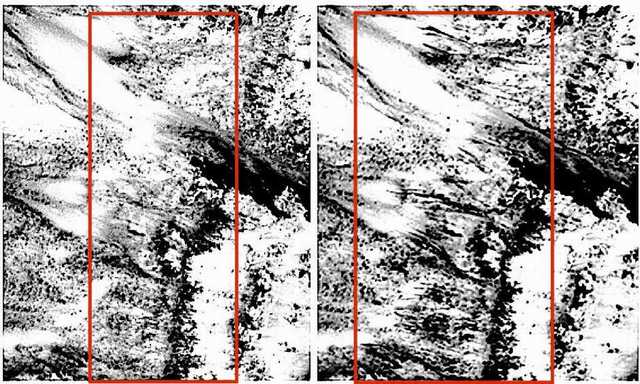
Figure 6. Mars HiRISE orbital photos (false colors desaturated). Left, Day 1. Right Day 34. Final length of fan-like specimens is 90 meters, with an estimated growth rate of 5 to 10 meters per day.

Figure 7. Sol 1143 documents area of soil devoid of spherical structures (circled in red). Seven days later, at least 18 spherical specimens have appeared in that same area.

Figure 8. Sol 1145 vs Sol 1148. Nine spherical and semi-spherical specimens lay upon the coarse grain sand of Meridiani Planum. Three days later, 12 additional spheres and semi-spheres appear and the original 9 have increases in size and diameter. Specimens are approximately 3-8 mm in size. Photographed by rover Opportunity.
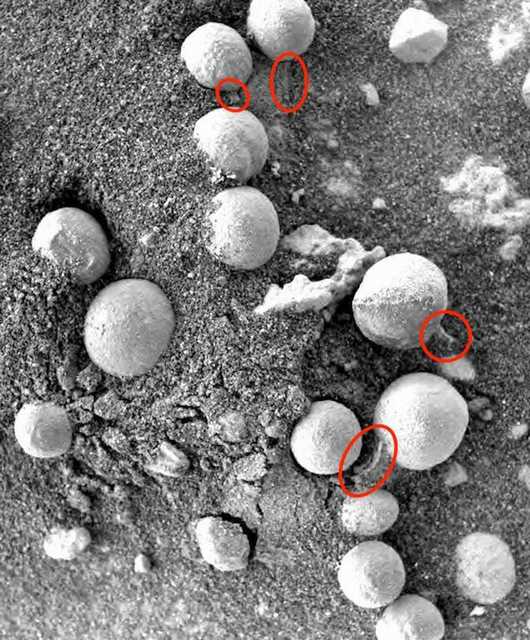
Figure 9. Sol 147. Photographed by Opportunity. Martian specimens approximately 3-8 mm in size resembling Puffballs (Basidiomycota), some with stalks or shedding white spore-like material (leprose).
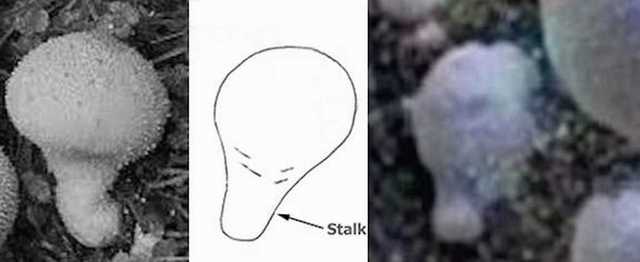
Figure 10. (Left) Terrestrial “puffball.” (Center) Diagram of “stalk.” (Right) Sol 257: Martian specimen.

Figure 11. Sol 257 photographed by Opportunity. Martian specimens approximately 3-8 mm in size resembling Puffballs (Basidiomycota), some with stalks or shedding white spore-like material (leprose) and lower cup, and universal veil that covers embryonic fungi (crustose).
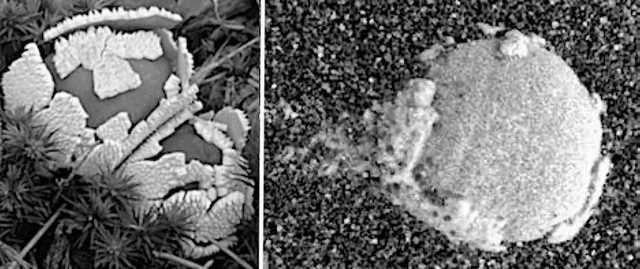
Figure 12. (Left) Terrestrial puffball shedding crustose. (Right) Sol 257: Shedding the universal veil that covers puffballs (crustose)?

Figure 13. (Left) Terrestrial puffball. (Right) Sol 147. specimens resembling puffballs.
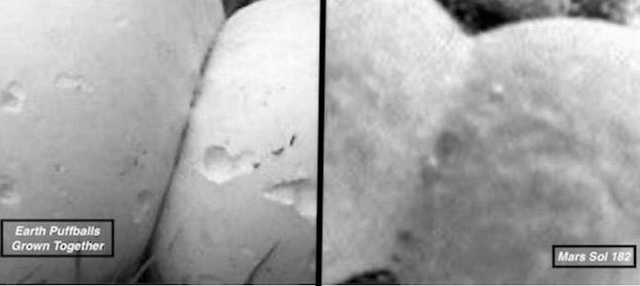
Figure 14. (Left) Terrestrial puffball that have grown together. (Right) Sol 182 spherical specimens Mars that have “grown” together. Note membraneous fibrous tissue within the interconnecting callosums.
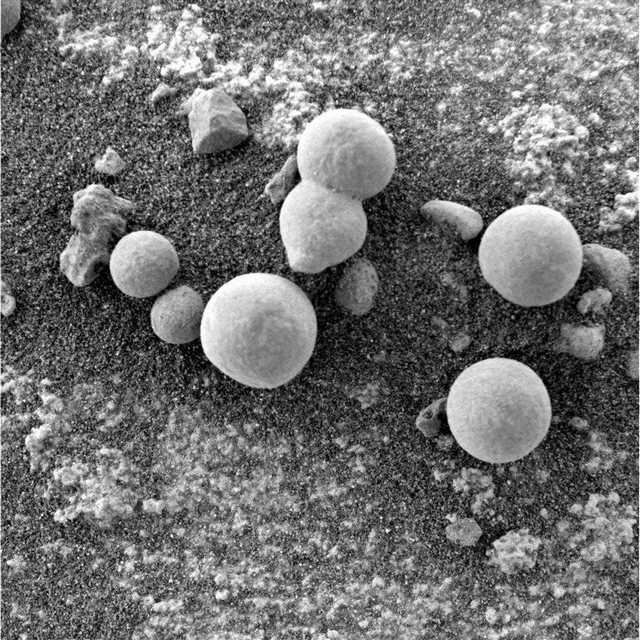
Figure 15. Sol 182 photographed by NASA Rover Opportunity. A majority of experts identified these specimens as "fungi" and "puffballs" (Joseph 2016). Note what appears to be white powder-chunky-spore-like material (leprose) littering the surface (see Figure 16 for a 200% magnification of the white material lying upon the surface). Spherical specimens are approximately 3-8 mm in size.

Figure 16. Sol 182. A 200% magnification of white powder-chunky-spore-leprose-like material littering the surface as seen in Figure 15. Note numerous (embryonic?) open-tubular- and stemmed-mushroom-shapes.
2. Leprose, Crustose, Puffball Fungal Spores and Fruiting Bodies?
Puffball leprose begins as crustose, the outer skin (universal veil) which eventually begins to break apart, falls to the surrounding surface and disintegrates (Petersen 2013; Roberts & Evans 2011). Puffballs generally do not have spore producing gills or pores. Instead, they produce fluffy white spores from specialized cells called basidia (Petersen 2013; Roberts & Evans 2011). These white spores, along with leprose, will litter the surface which, in combination can be powdery as well as rough, scabrous, and granular (i.e. “leprose”). Hematite has no outer skin and does not produce spores, crustose or leprose.
As documented in this report, these Martian spherical specimens are often surrounded by rough, scabrous, granular, flaky, fluffy white substances. Moreover an examination of Figure 16, reveals, at 200% magnification, the presence of numerous open-tubular- and stemmed-mushroom-shapes enmeshed within the white fluffy scabrous material.
Puffballs may also have stalks (stems), that become root-like rhizoids and subsurface mycelia that absorb nutrients. Fungal mycelia also grow into visible, above-ground, spore-bearing reproductive mushroom-shaped and spherical structures, called fruiting bodies (Petersen 2013; Roberts & Evans 2011) at which point they may become rigid and seemingly calcified.
3. Waxing, Waning, Shape Shifting, Movement and Dimpled Martian Mushrooms?
Rapid waxing and waning appears to be a common feature of biological-like specimens photographed on Mars. For example, and as reported here: on Sol 173, the rover Curiosity photographed numerous white colored amorphous specimens attached within a Martian rock shelter (Figure 18). Several of these specimens have a bowl- shaped appearance. Thirteen days later, as photographed on Mars day Sol 17 (Figure 19), the specimens have multiplied and increased in size. Yet another white amorphous specimen depicted in Sol 173, has almost completely disappeared when photographed 19 days (sol) later on Sol 192 (Figures 20-21).
A white amorphous mass, with appendages, appears in a rock shelter on Sol 528. However, it changes shape and a portion moves into a new location, as photographed the following day (Sol 529). When photographed on Sol 530, this amorphous mass has almost completely disappeared from its original location. A small portion of the remainder appears to have moved to a new location on the opposite inner-wall of the crevice (Figure 17). If this is fungus, slime mould, or a slime fungus, is completely unknown. However, amorphous shape shifting, alterations in mass and movement from one location to another, are typical of slime moulds and indication of behavior and life.
A comparison of Sol 192 and Sol 270, shows an increase in the mass of networks of white strands that resemble plasmodium, bulbous fruiting sporangia and interconnected clumps, when photographed months later (Figure 22). Photographs of additional surface features in this area on Sol 270, reveals the presence of dimpled donut-shaped specimens that resemble lichen genus Ochrolechia, or possibly Russula and “milkcap” mushrooms e.g. Lactarius quietus; Lactarius torminosus; Lactarius deliciosus (as described by Petersen 2013; Roberts & Evans 2011). These mushroom-like features surround these networks of what appear be rigid, encrusted and seemingly calcified, plasmodium and sporangia (Figures 22-27). Some of these donut-shaped specimens also appear to turn white and are linked to these tangled networks of white strands some of which lift up above the soil, casting shadows, and are connected to what resemble fruiting bodies (Figure 23). To speculate, perhaps these white tangled “veins” represent fungal hyphae or fossilized plasmodium and sporangia: networks of fruiting bodies?

Figure 17. Sols 528, 529, 530. (Gale Crater) White amorphous mass alters shape, location, and almost completely disappears from inside the crevice of a rock shelter over a three day (sol) period. Central mass is approximately 6 mm in diameter on Sol 528 and a thin tendril of white appears extends downward and another upward along the lip of the crevice. On 529 the lower tendril has disappeared and there are two tendrils snaking upward. On 530 the white mass and tendrils have nearly disappeared.

Figure 18. Sol 173 (Gale Crater). Compare white specimens (each approximately 3-6 mm in size) with Sol 186.

Figure 19. Sol 186 (Gale Crater). White specimens increase in mass and number as compared to Sol 173 (Figure 18).
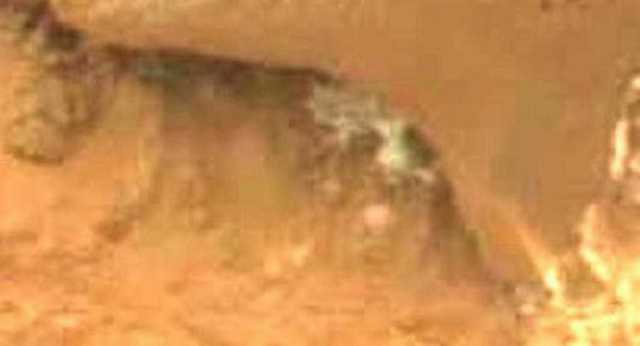
Figure 20. Sol 173. (Gale Crater) White amorphous masses from 3 to 5 mm in size, attached to underside of the lip of the rock shelter. Nineteen days later, the amorphous mass has disappeared (see Figure 19. Below).
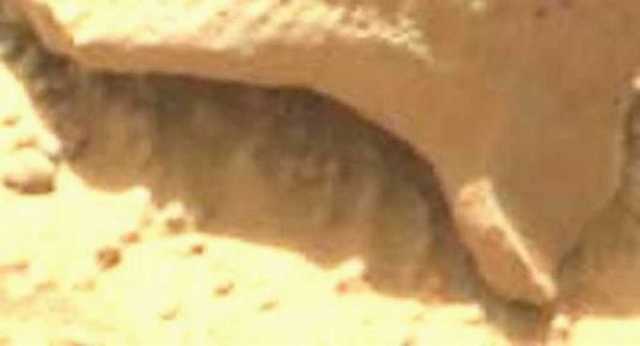
Figure 21. Sol 192. (Gale Crater) White amorphous mass has disappeared from underside of the lip of the rock shelter (compare with Figure 20, Sol 173).

Figure 22. Sol 192 vs Sol 270. Increase in mass of white amorphous networks after 78 days (sols). Note water pathways and dimpled-donut shaped specimens in upper panels. Note water pathway.
4. Minerals, Mushrooms, Fossilized Protoplasmic Plasmodium Sporangia?
Encrusted interconnected networks of white strands punctuated with white clumps and often surrounded by dimpled donut-shaped mushroom-like formations can be discerned in Figures 22-27. If the white specimens are abiogenic and consist of gypsum, calcium, or quartz, of if they are calcified fungal mycelium or encrusted plasmodium and protoplasmic tendrils punctuated with fossilized bulbous fruiting bodies (sporangia) is unknown. By contrast, there are no abiogenic analogs for the numerous dimpled-donut-shaped specimens which instead resemble the lichen genus Ochrolechia, and terrestrial “milkcap mushrooms” (e.g. Lactarius quietus; Lactarius torminosus; Lactarius deliciosus) and the “donut” shaped Russulas (Petersen 2013; Roberts & Evans 2011), and less so dimpled lichens which have been previously tentatively identified in Gale Crater (Joseph et al. 2020c,d). There is also evidence of water pathways (Figures 22, 24, 25) which would provide moisture to living organisms.
Terrestrial dimpled mushrooms commonly secrete a milky substance that can have the consistency of latex (Petersen 2013; Roberts & Evans 2011). Might this explain the solidified appearance of these strands, or why some of the white encrusted material is entangled with dimpled-bulbous formations? One can only speculate.
It is possible these specimens consist of minerals or other non-biological substances. For example, gypsum and quartz may be abundant in Gale Crater (Nachon et al. 2014; McLennan et al. 2013; Rapin et al. 2016). However, if these tangled webs of strands consist of quartz or gypsum this does not rule out biology. For example, algae (Cyanobacteria) are often found in association with gypsum which is a favored substrate for extremophile algae/cyanobacteria (Bothe, 2019). The green alga Closterium (and its relatives), also accumulates gypsum and stores it as crystals (Sr and/or Ba sulfates) in vacuoles located in the apices of their spindle-shaped cells (He et al. 2014).
Alternatively, the white-colored “veins” may consist of quartz and hydrated calcium sulfates which have been identified in the Gale Crater where these specimens were photographed (Nachon et al. 2014; McLennan et al. 2013). Quartz is associated with biological activity, and quartz, dominated by lichens have been identified (Jung, et al. 2019). Furthermore, cyanobacteria, green algae and microfungi will colonize the inner structures of small quartz gravel (Jung, et al. 2019). Another possibility is calcium carbonate--byproduct of cyanobacterial activity. Calcium carbonate is precipitated in the mucous of algae (cyanobacteria) via photosynthetic CO2 or HCO3- uptake (Devaud and Girardclos, 2001).
These white “veins” punctuated with white clumps, were photographed in association with formations that have a central depression or nuclei giving them a "donut shape" characteristics of fungal mushrooms and the lichen genus Ochrolechia. Given that strands of these white “veins” are above ground and sometimes rise above the surface and cast a shadow beneath them (Figure 23), then the most likely explanation is that these networks have a biological origin have become encrusted and rigid or photo-bleached due to surface conditions and radiation exposure. Again, one can only speculate.

Figure 23. Strands that snake across and sometimes rise above the surface. If biological, these specimens may consist of calcified fungal mycelium or encrusted plasmodium and protoplasmic tendrils punctuated with fossilized bulbous fruiting bodies (sporangia). Or they may be very unusual abiogenic formations.

Figure 24. Sol 270 Donut shaped specimens--approximately 1-2 mm in size--resembling the lichen genus Ochrolechia, and possibly “milkcap mushrooms” and Russulas. The white specimens resemble plasmodium, the bulbous fruiting sporangia and interconnected clumps of biological material that may have become rigid, encrusted and seemingly calcified. Or these networking strands may be very unusual abiogenic formations consisting of quartz, calcium, or gypsum. Note water pathway.

Figure 25. Sol 270 Donut-shaped specimens --approximately 1-2 mm in size-- adjacent to networks that resemble plasmodium and bulbous fruiting sporangia. If biological these, interconnected clumps of material may have become rigid, encrusted, calcified or fossilized. Or these networking strands may be very unusual abiogenic formations consisting of quartz, calcium, or gypsum. Note water pathway.
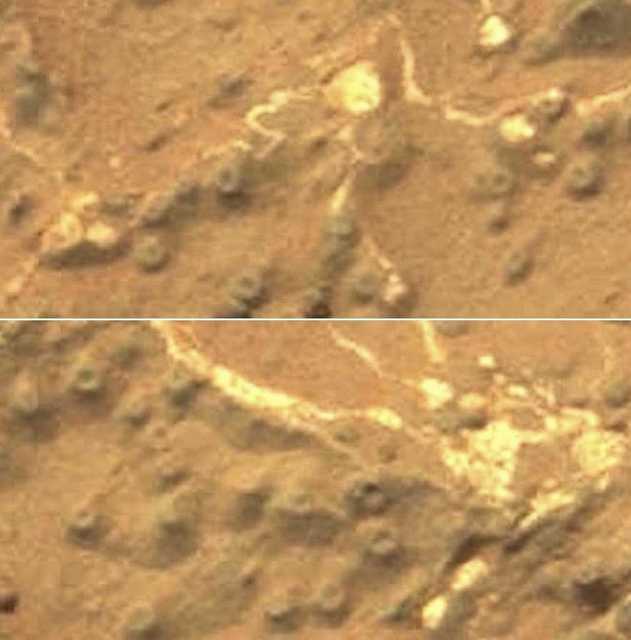
Figure 26. Sol 270 Donut shaped specimens --approximately 1-2 mm in size-- resembling the lichen genus Ochrolechia, and “milkcap mushrooms” (e.g. Lactarius quietus; Lactarius torminosus; Lactarius deliciosus) and the “donut” shaped Russulas. The white specimens resemble plasmodium, bulbous fruiting sporangia and interconnected clumps of material--all of which may have become rigid, encrusted, calcified when exposed to surface conditions. Or these networking strands may be very unusual abiogenic formations consisting of quartz, calcium, or gypsum.

Figure 27. Sol 270 Donut shaped specimens --approximately 1mm in size--resembling the lichen genus Ochrolechia, and terrestrial “milkcap mushrooms” (e.g. Lactarius quietus; Lactarius torminosus; Lactarius deliciosus) and some species of Russula.
5. Pareidolia and the Denial of Diverse Evidence of Martian Life
Nearly three dozen scientists with a history of major peer reviewed scientific accomplishment have found evidence indicative of Martian biology (Baucon et al. 2020; DiGregorio 2018; Bianciardi et al. 2014; Dass, 2017; Joseph et al. 2019, 2020a,b,c,d,e,f; Kaźmierczak 2016, 2020; Levin & Straat, 1977, 1977, 2016; Levin et al. 1978; McKay et al. 2009; Noffke 2015; Rizzo 2020; Rizzo & Cantasano 2009, 2016; Ruffi & Farmer, 2016; Small 2015; Thomas - Keprta et al. 2009). Diverse, putative, biological specimens resembling lichens, fungi, algae and open cone apertures for the venting of oxygen produced via photosynthesis have also been photographed in close proximity to one another (Joseph et al. 2020a,b).
However, these and other reports of biological specimens have been ignored by NASA and dismissed as “illusions” or a form of pareidolia (Geller 2014; Plait 2012; see discussions by Levin & Straat, 2018; Rizzo & Cantansano, 2016; Rowan, 2016). Instead, these life-like specimens are attributed entirely to minerals and abiogenic geological activity including wind, weathering, mud volcanoes, geysers and sublimating ice and melting carbon dioxide (Dundas et al. 2015; Hansen et al. 2010; Kieffer et al. 2006; Pilorget 2011; Portyankina et al. 2020; Soderblomet et al. 2004; Squires et al. 2004).
Conversely, it has been argued that those promoting abiotic explanations are also suffering from “pareidolia” (Joseph 2014b; see also Rizzo & Cantansano, 2016) a consequence of group and institutional peer pressure to enforce conformity of thought (Asch, 1955, 1956; Joseph 2014b; Levin & Straat, 2018; Rowan, 2016): as best illustrated by the parable of the “Emperor’s New Suit.” As demonstrated experimentally, peer pressure and the need to conform are so powerful that those affected are unable to accurately see what is right before their eyes (Asch, 1951, 1952, 1955, 1956).
6. Hematite and Speculation Based on Pictures: NASA and Planetary Geologists Rely on Photos.
A common complaint is that those advocating in favor of biology are making their judgments based on the analysis of pictures. There is truth to this criticism. However, Rizzo (2020) has discussed, at length, the fact that NASA’s and other planetary geologists commonly base their interpretations of Martian features, on pictures, not factual data. Case in point: based merely on orbital photos, NASA (2016) argues that the massive “araneiforms” which appear every Spring and wane every Autumn and Winter most likely “result from thawing of seasonal carbon-dioxide ice” --better known as "dry ice”--even though Carbon-dioxide ice is a semi-translucent white and not black and there are no Earth analogs to support what is nothing more than speculation. The only “araneiforms” analogs from Earth are biological; and yet this possibility is completely ignored except by a few scientists (Ganti et al. 2003; Joseph et al. 2020c; Kereszturi et al. 2012; Ness 2001).
The repeated claim that the Martian spheres are hematite concretions is another example of speculation, and which has been criticized by Burt and others (Burt et al. 2005; Knauth et al; 2005; Joseph et al. 2020a,b). Hematite is not gray or white in color (unless submerged in a standing body of water) and does not grow out of the ground, and expand in size, in just three to seven days (Figures 6-8, 31-33). Nor does hematite grow skyward, in huge colonies, attached to rocks by stalks topped by bulbous mushroom-shaped caps (Joseph et al 2019, 2020a,b). These latter Martian specimens closely resemble colonies of photosynthesizing lichens (see Figure 28-30); the existence of which would account for the seasonal increases in Martian atmospheric oxygen (Joseph et al. 2020a,b). Nor is hematite blue, purple or green ( Figure 30-32, 35) or easily broken and hollow inside (Figures 31-32, 34). Nevertheless, NASA and many planetary scientists embrace the hematite hypothesis and reject biological explanations.
A careful review of the methodology and instrumentation reported by the Opportunity team, reveals that the stalked-mushroom-shaped specimens and spheres atop the soil were never individually or selectively analyzed by Opportunity's suite of sampling instruments for hematite. Instead, individual samples, inferred to contain hematite, consisted of oblong rocks (see Figure 6 in Belle et al. 2004). Claims about hematite were also based on panoramic images, and claims about the averaging of high and low "temperatures" (Klingelhöfer et al. 2004) and the spectral signatures of false colors (Soderblom et al. 2004), with spectra selectively eliminated until what remained was interpreted as similar to the spectral signature of hematite photographed in a laboratory (Christensen et al. 2004), when the results were a "poor fit" for hematite and there were significant problems with calibration (Glotch and Banfield, 2006).
The Opportunity's cameras and instruments were not capable of taking true color photos. Although the true color of the landscape, outcrops, sand, dust, dirt, and rocks are unknown, composite false color images were generated by the Opportunity's panoramic camera's nanometer filters (Soderblom et al. 2004). Based on these "color composites" massive amounts of solid blues and greens were painted throughout the lower landscape--and if true colors, these blues and green would not be indicative of hematite, but pools of water and vast fields of chlorophyll-containing living organisms (Joseph et al. 2020b). Hematite is not green or blue. In fact, spectral signatures indicative of chlorophyll have been detected by the Pancam and Mars Science Laboratory (MSL) aboard the rover Curiosity. Stromberg and colleagues (2014) reported that features "associated with chlorophyll can be distinguished" and that the "chlorophyll spectral feature is observable in both Pancam and Mastcam" spectra obtained by the rover Curiosity's MSL in Gale Crater (see also Stromberg et al. 2019). Hematite is not green and does not contain chlorophyll. Moreover, based on false colors, the spherules of Eagle Crater were judged to be yellow, orange, and purple (Soderblom et al. 2004); colors not associated with hematite, but living organisms. Nor does terrestrial hematite have an outer coat that it sheds or a mushroom shape and stem and grow upward and outward from the tops of rocks or soil (Figures 7-8, 28-31).

Figure 28. Sol 85 photographed by the rover Opportunity. Example of vast colonies of lichen-like formations with stems, attached to rocks and jutting upward and topped by bulbous caps.

Figure 29. Sol 85 photographed by the rover Opportunity. Example of vast colonies of lichen-like formations with stems, attached to rocks and jutting upward and topped by bulbous caps.

Figure 30. Opportunity, False Colors by JPL/NASA. (Upper Left) Sol 88. Example of vast colonies of thousands of mushroom-lichen-shaped specimens, attached to rocks with thin stems topped with bulbous caps, similar to photosynthesizing organisms (see Figures Joseph et al. 2020a). (Upper Right) Sol 37 Examples of long stemmed, bulbous-mushroom-lichen-shaped specimens (Bottom) Sol 183. Vast fields of innumerable spherical specimens photographed in Meridian Planum.
7. Growth and Regrowth After Crushing by Rover Wheels: Hematite Refuted
Hematite is characteristically harder that iron and yet brittle and can be identified by the presence of a reddish streak. When struck repeatedly by hammers and other tools, pieces may break off and it may shatter into large jagged chunks. It can only be reduced to a red powder by extensive grinding.
Although hematite may be generated in volcanoes, when produced in hot springs it is in part fashioned via biological activity over hundreds of thousands of years (reviewed by Joseph et al. 2019). Hematite cannot spontaneously recombine, reform, regrow, emerge from the soil and increase in size in just three and seven days, or engage in other behaviors. These are the province and hallmarks of biology.
As documented in Figures 31-32 vast fields of spherical specimens were crushed, obliterated, disappear and/or are turned into blotchy smears by the wheels and treads of the rover Opportunity. However, in less than 650 days (Sols) innumerable spherical specimens re-appear atop the crests of old tire tracks; and many have mushroom-shapes with stems. These spheres, including those that emerged from the soil in 3 days (Figure 8) and 7 days (Figure 7), in all respects, have the characteristics, morphology and patterns of growth characteristic of living organisms. By contrast, claims that these spheres are hematite are based on speculation, inference, false colors, the averaging of temperatures, ad hoc calibration adjustments, and the selective removal of spectral signals generalized from panoramic images that included sand, soil, dust, oblong rocks, and outcrops (Bell et al. 2004; Christensen et al. 2004; Klingelhöfer et al. 2004; Rieder et al. 2004; Squires et al. 2004). The Opportunity team’s instruments were not even mineral specific. The evidence favors biology, not mineralogy.

Figure 31. (Left) Opportunity, Sol 1232. Crushing, obliteration of sphericals. (Right) Sol 1319: Note
sphericals appear in the crests of old tire tracks indicating they were not blown or knocked into place.

Figure 32. On Sol 952, the rover wheels obliterated almost all spheres. By contrast, numerous spherical specimens, many with stems and mushroom shapes have grown into the old tire tracks formed on Sol 336. These “new” spheres are located predominantly on top of the the crests (see also Figure 31).

Figure 33. Photographed on Sol 952, segment from Figure 32. Growth of spherical and stemmed-mushroom-shaped specimens that appeared 616 days (sol) after the rover Opportunity wheels obliterated all evidence of these spheres on Sol 336.

Figure 34. The spheres upon the surface are fragile, easily broken and will leak and shed their interiors when broken.

Figure 35. Photographed by rover Opportunity. Vast fields of spherical specimens, many with stalks above the surface. The “greenish” color may indicate chlorophyll.
A Summary of Evidence Favoring Martian Life
The question of if there is, or was, life on Mars, is extremely controversial. Even those favoring biology have not yet claimed to have found definitive proof. Admittedly: similarities in morphology are not proof of life, even when the evidence includes a wide range of specimens that appear biological including: putative fossilized stromatolites, microbial mats, tube worms, metazoans, and calcareous algae (Baucon et al 2020; Bianciardi et al 2014; Joseph et al 2020a,d-g; Kazmierczak 2016, 2020; Noffke 2015; Rabb, 2018; Rizzo et al. 2020; Ruff & Farmer 2016; Small 2015). Then there is the biological residue discovered in Martian meteorites (Thomas-Keprta, et al. 2009; McKay et al. 2009); and what appear to be living organisms including algae, fungi, and lichens (Dass 2017, Joseph 2014a 2016; Joseph et al. 2019, Joseph et al 2020a,b; Krupa 2017; Levin et al. 1978; Levin & Straat 2016; Rabb 2015, 2018, Small 2015); and biological activity detected in Martian soil samples (Levin & Straat 1976, 1977; 2016); and the 9 spherical specimens that increased in size and the 12 that emerged from beneath the soil in just three days (Sols 1145 and 1148) and seven days (Sol 1143 and 1150); and spheres and stemmed-mushroom shapes which emerge in old tire tracks formed by the rover Opportunity.
Massive black araneiforms emerge in the Spring, grow rapidly, and then rapidly wane in the Autumn and Winter only to re-emerge the following Spring. This evidence may represent the growth of massive colonies of black fungi, lichens, algae, methanogens, and sulfur reducing organisms (Ganti et al. 2003; Ness, 2001; Joseph et al. 2020c; Kereszturi et al. 2012). In support of this hypothesis are sequential images (Figures 36,37) of what may be black fungal bacterial growth on the rovers; but which may also be living contaminants transferred from Earth (Joseph et al. 2019s, 2020a).

Figure 36. Mars Sol 2718 (upper left) vs Sol 2813 (right and bottom)--Growth of specimens resembling bacteria and fungi on the Rover, Opportunity, after 95 (Martian) days on Mars--

Figure 37. Sol 51 (left) vs Sol 1089 (right). A portion of the Chem Cam deck of the Rover Curiosity Photographed on Sol 51 and 1038 Martian days later (Sol 1089). Possible evidence of the growth of fungi and bacteria. As previously detailed, these specimens may represent the flourishing of living contaminants which adhered to this equipment and survived the transfer from Earth to Mars (Joseph et al. 2019).

Figure 38. Top (Sol 52). Rover Curosity: Mastcam photo of the interior, flooring, and shelter of an exposed compartment after 51 Martian days/Sols. Bottom (Sol 1089). Possible fungal contamination or bio-corrosion of the interior, walls, and flooring of an exposed compartment in the chem cam desk after 1089 days/Sols. Another possibility: Salt which came to adhere only to the interior of the surface perhaps secondary to moisture? Or perhaps fungi may have been transported to Mars already attached to the rover (Joseph et al. 2019).

Figure 39. (Left / Sol 1089) Contamination of open compartment of the rover with (Right Sol 1162) white amorphous mass. If a fungus, then it is possible the rover may have been contaminated when this specimen was photographed 27 days/Sols earlier.
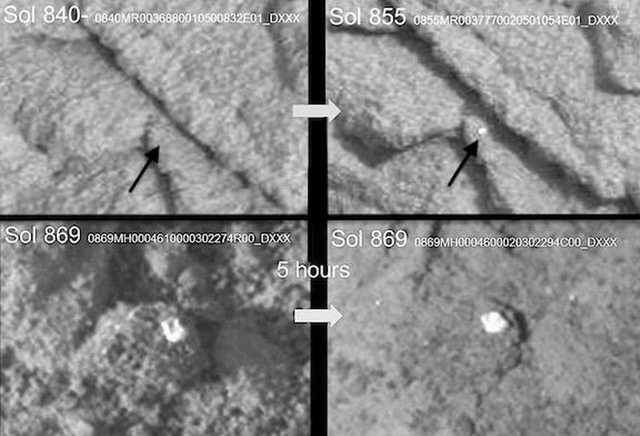
Figure 40.. Photographed by rover Curiosity. Top Row: Sol 840-arrow indicates gray mm-size dimpled-spoked formations. Fifteen days later (Sol 855) a white colored formation appears. Bottom Row: Square-shaped-specimen near shadows with vertical “side-walls” and square-concave center (Sol 869). Five hours later (Sol 969) in response to full sunlight it’s become oval in shape and “side-walls” are lateral.
In this report we now provide additional evidence of waxing, waning, and the multiplication of amorphous specimens, some of which appear to move to different locations; as well what may be networks of mycelium, plasmodium, bulbous fruiting sporangia that have become rigid, encrusted, calcified or fossilized. Moreover, these networks are adjacent to dimpled-donut-shaped specimens that resemble the lichen genus Ochrolechia, and “milkcap mushrooms” and some species of Russula. Rapid waxing and waning in size appears to be a common attribute of a wide variety of Martian specimens.
Waxing vs waning of amorphous and fungal-like spherical specimens have been photographed in the equatorial regions at ground level (Figures 6, 17-22). The huge black and growing araneiforms have been photographed from orbit, in the arctic regions (Figures 1-6,45-51). However, until this report, there have been no attempts to statistically analyze and quantify what appears to be growth. The purpose of this study, therefore, was to perform a statistical analysis and quantitatively compare sequential images to determine if specimens (A) resembling fungal “puffballs” are growing. If biological, it can be predicted that as they increase in diameter and size, they should become closer together and the distance between them should decrease. By contrast, if due to wind, they should not increase in diameter and the distance between them should not decrease but remain the same or randomly increase and decrease as they are moved by wind. Likewise, if the (B) araneiforms show statistically significant evidence of parallel expansion over time that could be indicative of life.
Experiment 1
Method
Images: Two images were analysed from Mars, viz. Opportunity Sol 1145 and Sol 1148 (Figures 8, 41). Various spheres are visible and those clearly present in the two images have been labelled A to I.
Theory: The problem in making quantitative comparisons between Sols is that there may be variations in the exact positioning of the camera on the different occasions. Hence, a method of determining growth/expansion was devised based on relative measures of ratios and is based on the premise that if growth has occurred then the distance between the adjacent edges (E) of two mushrooms would reduce while the distance between their centres (C) would remain more or less constant. Consequently, the ratio of the centre distances (C-C) to edge distances (E-E) would increase on sol 1148 if expansion/growth has occurred in the intervening period.
Image analysis: Images were analyzed using ‘Image J' software developed by the National Institute of Health (NIH), Bethesda, USA (Armstrong and Bradwell 2010, Rizzo et al 2021). Each image was opened using the software and magnified to clearly reveal the structures of interest. Images were manipulated using brightness, contrast, sharpening, and if necessary edge detection, to optimize the appearance of the mushrooms. The following protocol was used to estimate whether there growth/ expansion of the mushrooms had occurred between Sols (Figures 8, 41-44): (1) a circle of best fit was fitted to each of the profiles A to I, (2) the centre of each circle was determined by drawing three ‘chords’ at different locations on each circle, the points of intersection of the lines drawn at the midpoint of each chord and normal to it define the centre, (3) for each pair of adjacent mushrooms, the distance between their centres (C-C) and edges (E-E) was measured, (4) for each pair of adjacent mushrooms, the ratio C-C/E-E was calculated. Nine pairs of mushrooms visible in Figures 8, 41 were analysed. The average change in the C-C/E-E ratio on sols 1145 and 1148 was also compared using a paired sample ‘t’ test. An applied cluster analysis (the paired group method, using the Euclidean index of similarity) was then performed using the PAST statistical package (Hammer et al., 2001) to distinguish and determine changing ratios between the existing groups within derived data (group 1: E-F and F-G; group 2: E-G, A-B, A-I and B-C; group 3: B-I, A-C and D-E). As measure from the center points, a statistically significant change in the ratios and distances between the puffballs was confirmed; i.e. all groupings became closer together due to specimen diameter expansion.
Results and Discussion: Growth of Martian Mushrooms?
Examination of Figures 41 reveals 9 puffball-like structures on sol 1145 (A to I). The same structures are visible on sol 1148 together with 13 new structures. A comparison of the C-C/E-E ratios for 9 pairings of mushrooms are shown in Table 1 and Figures 41-44). The C-C/E-E ratio increased from Sol 1145 to Sol 1148 for each of the 9 comparisons of pairs of spheres, the greatest change in ratio being observed for comparison of distances between spheres E,F and the smallest between spheres B,I. A paired sample ‘t’ test gave a value of ‘t’ = 3.28 (8DF, P = 0.011) indicating a significant increase in the C-C/E-E ratio from sol 1145 to 1148; the probability of a chance effect being approximately 1 in 100.
On Sol 1148, the existing puffballs increased in size and are closer together indicating growth and expansion, as confirmed by statistical and cluster analysis. To summarized, the larger the difference in ratios between sol 1145 and 1148 in table 1 indicates the specimens grew closer together. EF and AI most and DE and BI the least. The cluster analysis emphasizes that pairs of spheres behave differently with three groups of responses: (1) EF and FG grew significantly closer and were the closest together on sol 1145 (2). EG, AB, AI, BC grew significantly closer but were further apart on sol 1145. (3) BI, AC, DE which grow the least. These impressions are further confirmed by an examination of Figures 41-44.
It is important to stress that the statistical method is based in relative measures and makes no assumptions about the positioning of the camera. Second, there is no evidence of significant wind effects at the site between Sols 1145 and 1148 (Joseph et al. 2020b). Third, there are distinctive features at the edges of the spheres (e.g. A and G) they appear on the perimeter of the structure on both Sols and therefore, to have moved laterally. Fourth these specimens appear fragile (see Figures 32-33) and they are easily crushed. When the RAT was deployed on sol 1168, all of the spherical specimens were crushed leaving only a crusty-fluffy white debris. Therefore, it is highly probable that the data truly indicates significant degrees of lateral ‘growth’ of the these spherical specimens between sol 1145 an 1148.

Figure 41. Comparison of Sol 145 (Left) and Sol 148 (Right). Specimens at top (A,b,C,I) have not moved in relation to one another as measured from the center of the specimens. Instead they have expanded in size and became significantly closer together. Specimens at bottom (D,E,F,G) have increased in size, expanded closer together, but also moved in relation to one another, with specimen G show the greatest degree of positional change and movement which may account for the crescent hole (see specimens lower right).

Figure 42. Comparison of Sol 145 (Left) and Sol 148 (Right). As measured from their outer boundaries (top) and center (bottom) ABCI have grown closer together, and DEFG have move in relationship to one anther as measured from their centers; with D and G showing the most obvious changes in position.
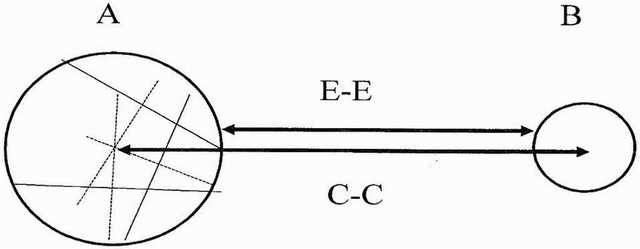
Figure 43. Method of measuring possible growth/expansion’ of fungal puffballs from Sol 1145 to sol 1148 (A, B adjacent ‘mushrooms, E-E = edge to edge distance, C-C = Centre to centre distance. Chords drawn in A are used to establish the centre of the circle.
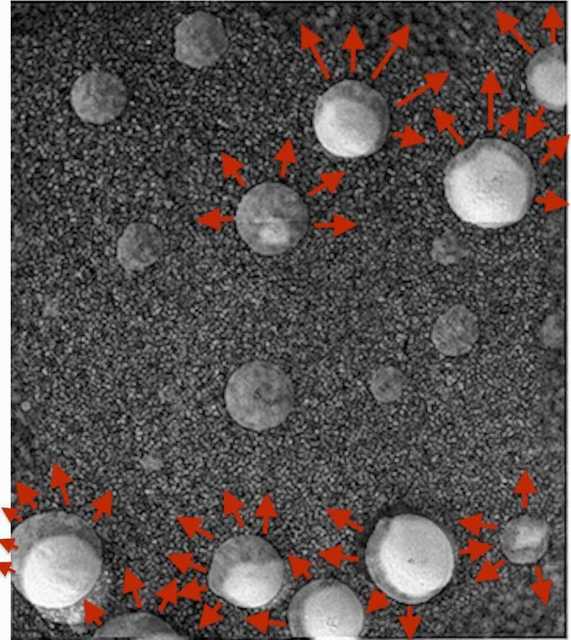
Figure 44. Clone of Sol 148 (at 50% opacity) placed on top of Sol 148, using center of sphere at top-center-right as anchor point. Patterns of growth and expansion. Specimens at the top expanded upward and to the right. Center most specimen expanded upward and to the right and left. Specimens at the bottom expanded upward and predominantly to the left. Sphere at lower left show greatest degree of positional change (moving up-leftward).
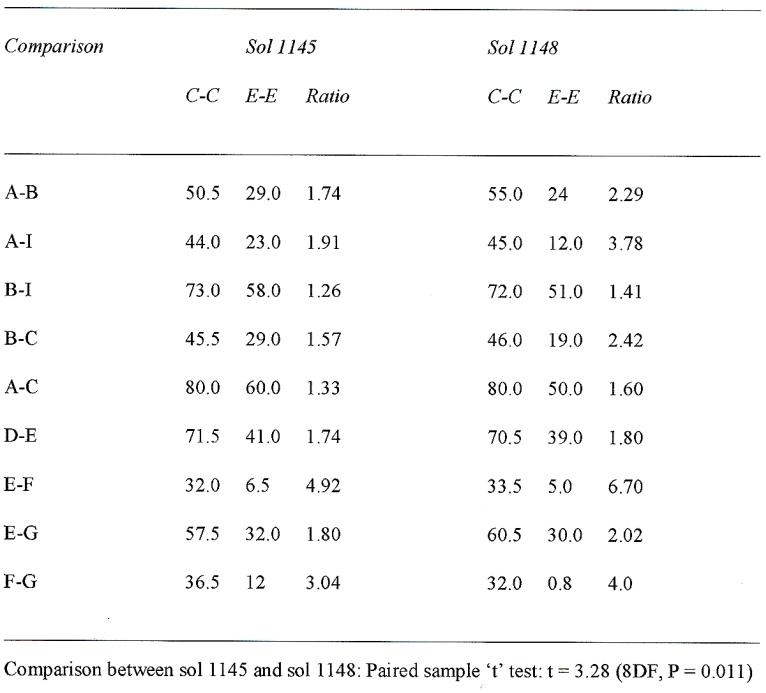
Table 1. Quantitative analysis of “Martian Mushrooms” (Puffballs) depicted in Figures 6, 38, on Sols 1145, 1148. Distances are measured in arbitrary units.
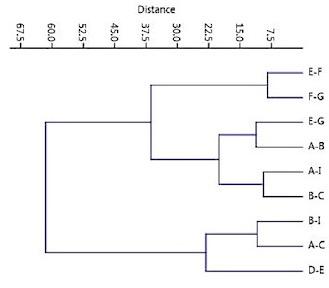
Table 2: Cluster Analysis: Classification of changes in Euclidean distance. Distinct groups expanded and became significantly closer together.
EXPERIMENT 2
A number of Martian features photographed by orbital satellite exhibit expansion or growth in sequential photographs. A notable example is the ‘black streaks’ which appear to gradually extend down the face of a dune (Figures 5, 47). The objective of this analysis was to compare the morphology of the ‘black streaks’ (Figure 5) on three occasions (Days 1, 22, and 35) to determine if there is any evidence of significant growth/expansion and therefore if these features could be evidence of life.
Method
Images: Three images taken on three days were analysed from Mars via orbital satellite (Figure 5). Various clusters of ‘black streaks’ are clearly visible and appear to expand over time apparently extending down the slope of a dune from its crest.
Image analysis: Images were analyzed using ‘Image J' software developed by the National Institute of Health (NIH), Bethesda, USA (Armstrong and Bradwell, 2010). Each image was manipulated using brightness, contrast, sharpening, and if necessary edge detection, to optimize the appearance of the streaks. The following metrics were recorded from the three images (Figure 5): (1) the number of individual streaks resolved by the image analysis (N), (2) the length of each streak from the crest down the slope (Ls), (3) the length of the upslope (Lds) from the bottom to the crest at the location of each streak, (4) the ratio of streak length up th slope length (Ls/Lds), (5) the width of each cluster of streaks at the midpoint of the longest streak within the cluster (W), and (6) the ratio of mean length of streaks per cluster to the width of the cluster (L/W). Comparison of metrics on the three occasions was made using analysis of variance (ANOVA) followed by Tukey’s Honestly Significant Difference (HSD) post-hoc test.
Results and Discussion
The data are summarized in Tables 2 and 3. First, there is a small increase in the number of streaks present that are resolved by the image analysis system from 11 on day 1 to 15 on day 34. Second, there are no significant differences in Lds among the sequential images (F = 0.86, P = 0.43), indicating that successive images were taken by the HiRISE satellite at approximately the same height. Third, mean Ls varied significantly among days (F = 6.42, P = 0.004), with post-hoc tests indicating that most of the increase in length occurred between day 1 and day 22--thus there is an initial, and significant growth spurt. Fourth, the Ls/Lds ratio also varied significantly among days (F 6.28,. P = 0.005), with the greatest increase between day 1 and day 22 with little significant change from day 22 to day 34--although it is apparent the the specimens increased in length. Fifth, the L/W ratio did not vary significantly among days (P > 0.05) indicating that length and width of the streaks increased in proportion from day 1 to day 22. In other words, these specimens, although separated by varying distance, and regardless of the terrain, all grew at the same rate--a feature of biology, not geology.
In addition to the appearance of some new streaks on days 22 and 34 which increased the width of the streak clusters, the data indicates an extension of the streaks up then down the slope over time from the crest. Most of this extension appears to occur between day 1 and day 22 with no significant change from day 22 to day 34. The statistical data confirms and indicates a significant degree of rapid parallel ‘growth’ between Sol 1 and 22.
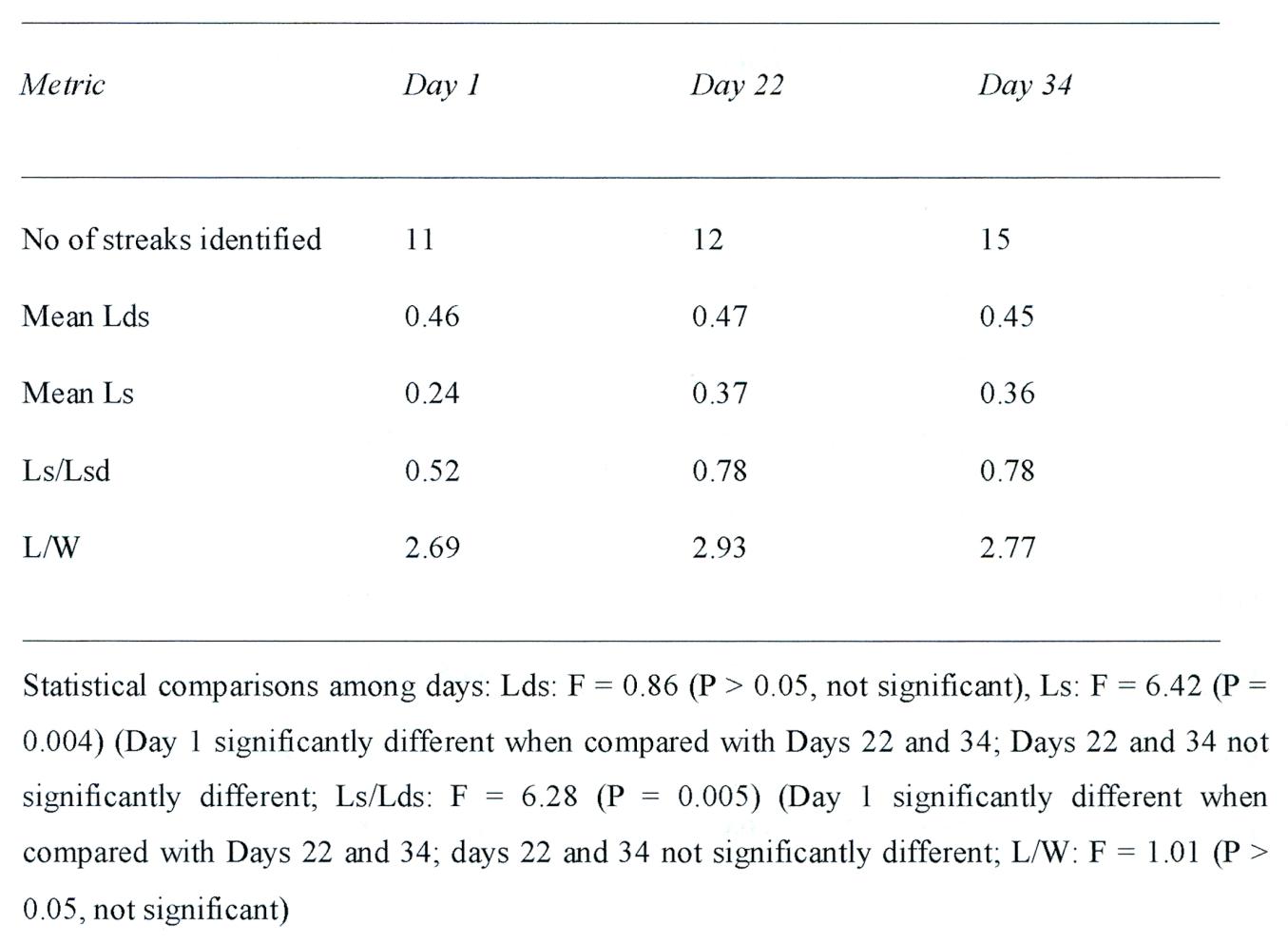
Table 3. Quantitative analysis of black streaks depicted in Figure 5 on three occasions by the HRISE satellite experiment. Absolute distances are measured in arbitrary units. Abbreviations: Lds = Distance from crest to bottom of slope. Ls = length of streak. Lw = Ratio of mean length of streak per cluster to width of cluster.
DISCUSSION
Based on statistical analysis of select samples, there is significant evidence of and increase in diameter and lateral ‘growth’ of spherical specimens identified as fungal puffballs, between sol 1145 an 1148 such that as they increased in size they also grew closer together. Likewise the arctic specimens, although separated by varying distance, and regardless of the terrain, all grew at the same rate in parallel and in an upslope direction--a feature of biology, not geology.
Nearly three dozen investigators, many with a history of major scientific accomplishment, have previously presented what they believe to be evidence of current and past life on Mars (Baucon et al. 2020; DiGregorio, 2018; Bianciardi et al. 2014; Dass, 2017; Joseph et al. 2020a,d-g; Kaźmierczak 2016, 2020; Levin & Straat, 1976, 1977, 2016; Levin et al. 1978; McKay et al. 2009; Noffke 2015; Rabb, 2018; Rizzo 2020; Rizzo & Cantasano 2016; Ruff & Farmer 2016; Small 2015; Thomas - Keprta et al. 2009). In this report, we provide evidence of movement, relocation, alteration in shape, increases and reductions in mass, and statistically significant findings of growth. As documented here, Martian specimens emerge from beneath the soil, increase in diameter, and, in the arctic, reaching hundreds of meters in size in the Spring and early Summer only to disappear by Autumn and Winter; whereas those in equatorial rock shelters change shape, move to different locations, or almost completely disappear or increase in mass and number: “behavior” that is characteristic of life.
Martian Puffballs? Statistical Analysis Supports Biological Growth
The focus of these statistical analysis included 9 puffball shaped specimens that were photographed on Sol 1145. Three days later twelve additional puffball specimens emerged from beneath the soil whereas the original 9 appear to have increased in size (Figures 8, 38-40). As documented in this report, a comparative statistical analysis indicates that the original 11 specimens significantly increased in diameter, such that the distance between them decreased in parallel due to mutual expansion. Joseph and colleagues (2020a,b) ruled out the possibility that wind may have uncovered these specimens. Specifically, as reviewed by Joseph et al,. (2020b) ground level wind speeds between 40 to 70 m/h are required to move coarse grained soil on Mars, and no strong winds, dust clouds, dust devils, or other indications of strong winds were observed, photographed, or reported during those three days in this vicinity of Mars. Hence, there is no evidence these spherical specimens were uncovered or moved by wind. Nor does the Sol 1148 photograph show any evidence that the surface has been disturbed by wind, as there are no parallel lineaments, ripples, waves, crests, or build-up of soil on one side of the specimens as would be expected of a directional wind (Kidron and Zohar 2014; Kidron et al. 2017). The only evidence of soil displacement is the partial hole within and adjacent to one of the original 11 specimens (Figures 8, 41-44). Soil displacement in the shape of a partial crescent hole, is what would be expected of soil-dwelling organisms growing in size whereas wind would fill in that shallow crescent (Kidron and Zohar 2014; Kidron et al. 2017). Moreover, all specimens appear to be on the same plane, level to one another, as if they are emerging and growing atop the soil. If the additional 12 specimens were uncovered from beneath the soil, then why do they not appear much lower, at a lower level than the original 9? If due to extremely powerful winds, then why weren’t some of the specimens pushed further apart or into new locations? If these Martian specimens are alive, then they had to have grown as a phenomenal rate. However, rapid growth appears to be the norm as documented in this report.
Araneiforms: Statistically Significant Evidence of Growth
As is now well documented massive black araneiforms appear on the surface of the upper northern and lower southern hemispheres of Mars and may grow at a rate of 5 to 10 km per day and become several hundred meters in size, length, and/or diameter. The emergence and growth of these formations parallels the melting of the polar ice- caps which consist of varying levels of frozen carbon dioxide and considerable amounts of water-ice (Bierson et al. 2016; Foss et al. 2017; Phillips et al. 2001; Thomas et al. 2016). After increasing in width, length, and diameter most of these huge black formations disappear by late Summer and Winter (Hansen et al. 2010; Portyankina et al. 2020). The following Spring and over the following years similar patterns emerge in the same locations (Figures 1-6, 45-51). As documented in this report, the growth patterns were found to be statistically significant with adjacent specimens growing at the same initially rapid rate. Moreover, as can be see in Figures 4 and 6, these arctic specimens grow upslope and curve toward one another.
Based merely on photos, an in the absence of Earthy analogs or any attempt to determine the identity of these formation, NASA (2016) has argued that these massive “araneiforms” “result from thawing of seasonal carbon-dioxide ice.” However, frozen Carbon-dioxide ice is not black but semi-transparent white; and this is obvious as based on an examination of orbital data and photographs of the southern and northern arctic regions of Mars. Furthermore, when CO2 melts, it would not lay on the ground but rise into the atmosphere. Specifically, it's been hypothesized that melting-pressure-induced release of CO2 causes the ejection of black sand and rocks (Pilorget 2011; Portyankina et al. 2020). Hence, as warming temperatures induce the sublimation of carbon dioxide it becomes gas that bursts upward or in multiple directions carrying black dust and other dark colored debris which spreads across the surface which is also being liquified by melt-water. Kieffer et al. (2006) have also proposed “that the seasonal ice cap forms an impermeable, translucent slab of CO2 ice that sublimates from the base, building up high-pressure gas beneath the slab. This gas levitates the ice, which eventually ruptures, producing high-velocity CO vents that erupt sand-sized grains in jets to form the spots.” Hence, there 2must be an unending supply of black sand and rocks which remain hidden until sprayed outward every Spring by melting glaciers. If true, then: (a) where do these black rocks and sand come from, (b) why do they disappear in Autumn, and (c) where do they go? and (d) why don’t these black rocks and sand appears simultaneously and immediately instead of expanding outward exponentially in a statistically significant pattern of growth? And why can’t these blacks rocks and sand be seen beneath the translucent ice before it melts and again after it freezes?

Figure 45. HiRISE orbital photo. Arctic dune with repeating black formations. Expanding formations atop and within arctic Martian sand dunes supports the hypothesis they begin growing within melt-water atop the dune and take identical expanding shapes, whereas thickening spider-web-like masses and grass-blade like appendages develop within the anterooms where water would also be expected to pool.
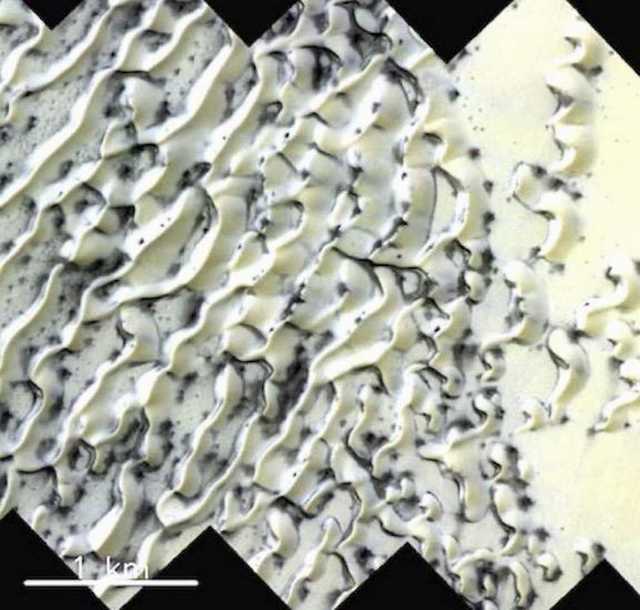
Figure 46. HiRISE orbital photo. Arctic Dunes, Mars. covered with black formations. Note repeating pattern.

Figure 47. HiRISE orbital photo. Arctic Dune, Mars. covered with black formations. Note repeating patterns.
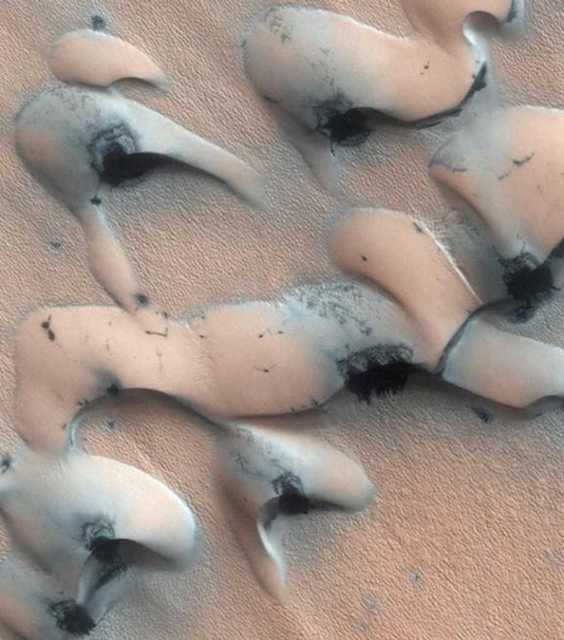
Figure 48. HiRISE. Arctic Dunes, Mars. covered with black formations. Note patterns repeat.

Figure 49. Mars: Orbital photos. Waning from black during the Summer and then gray in the Autumn/Winter. Photographed, via satellite, from orbit (From Ganti, et al. 2003).
Arctic Martian Melt Water and Growth of Dormant Organisms?
In the arctic similar formations assume identical expanding shapes and patterns of growth. Nearly identical formations appear on hundreds of dunes even those thousands of meters distant (Figures 45-51). These complex repeating patterns are characteristic of biology.
Mohlmann and Akos (2010) have proposed that brackish viscous meltwater is jetted onto the surface creating black streaks and other shapes. Brackish water and mud would nourish innumerable biological organisms (Joseph et al. 2020c) and could include “organic material, microbes, or even simple plants" as has been proposed by Ness (2001). Moreover, as arctic melt-water floods the surroundings black fungi, mould, lichens, algae, and other dormant organisms would begin to reproduce and proliferate in response to the availability of liquid and warming temperatures (Ganti et al. 2003; Ness, 2001; Joseph et al. 2020c; Kereszturi et al. 2012). Horvath and colleagues (Ganti et al. 2003; Kereszturi et al. 2012) have theorized that photosynthesizing organisms lay beneath, within, or upon the surface of arctic ice in a "dormant, desiccated state” and have identified algae / cyanobacteria (Kereszturi et al. 2012) and "lichens" (Ganti et al. 2003) as ideal candidates—lichens being composite algae-fungal organisms--all of which have been shown to survive direct exposure to space and Mars-like conditions (De Vera 2012, De Vera et al. 2019; De la Torre Noetzel et al. 2017, 2020). According to this scenario, these awaking photosynthesizing organisms might develop black pigmentation in response to radiation and sunlight, thereby absorbing energy and increasing localized warmth, and in so doing forming expanding repeating patterns and radial channels that grow larger and blacker (Joseph et al. 2020c).
Many terrestrial species of bacteria and fungi such as ‘black yeasts’ (Sterflinger 2006), and lichenized fungi (Armstrong 2013) and black fungi (Zakharova et al. 2014) develop dark or black pigments after emerging from dormancy. Algae/cyanobacteria, black fungi, and lichenized fungi dwelling in extremely frigid environments are often darkly pigmented and which enables them to absorb heat, and which via albedo, melts snow more rapidly thereby increasing the supply of water (Nordhagen 1928). Many species of lichens including members of the genus Peltigera are darkly colored and become almost black when wet and become grey when dry.
If biological and organic this would explain why these surface features change over time in response to warming, the availability of melt-water, and fade in response to cooling temperatures. Possibly, arctic fungi, mould, and immense colonies of dormant surface organisms, when exposed to melt water, become pigmented upon absorbing sunlight, flourish and rapidly reproduce, then dormant again when water evaporates and freezes in the Autumn and winter (Joseph et al. 2020c). This would explain why basically identical patterns repeat for years (Figure 50). Then there is the contribution of geysers and mud volcanoes that may spew a bacterial-infested organic soup onto the surface and providing nutrients. When waters evaporate and freeze, these colonies form spores, become dormant, migrate beneath the surface, or die and the dark coloration disappears; and then the pattern repeats the following Spring.
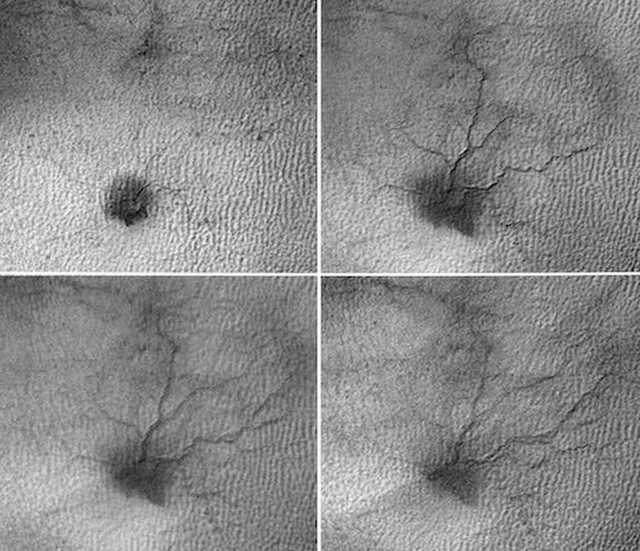
Figure 50. HiRise orbital photos of massive “spider-like” formation with tributaries, that repeat the same sequence over a 5 years time period: 8/5/2009 - 5/25/2015

Figure 51. Orbital photo of Mars. Martian geysers?
The Astrobiology of Martian Geysers and Mud Volcanoes
Investigators have also proposed that these massively growing black formations are produced by mud and dirty water expelled from mud volcanoes and geysers (reviewed by Joseph et al. 2020c). Nearly 20,000 circular cratered mounds believed to be similar to the "mud volcanoes" of Earth have been observed on Mars (Kumar et al. 2019; Pondrelli et al. 2011; Skinner & Adriano 2009).
These Martian "mud volcanoes" (MVs) commonly take the form of domes, cones, pits, and elliptical and circular mounds and may be found in clusters and belt-like chains (e.g., Dapremont and Wray 2020; Komatsu et al., 2016) with numerous pits that may be interconnected by subsurface “pipes” (Kumar, et al. 2019). The Martian MVs, therefore, appear to be fed by underground rivers, lakes, and reservoirs of water that are interconnected or adjacent to one another (Remizovschi et al. 2018; Pozzobon et al. 2019). This would explain why numerous spidery araneiforms, and what appears to be black geysers and MV appear adjacent to one another. This underground watery organic soup once sufficiently pressurized or heated (at temperatures as low as 2 °C (36 °F) will rush upward and puncture and pour forth upon the surface (Skinner & Adriano 2009; Remizovschi et al. 2018).
Like the mud volcanoes of Earth, it is not likely that Martian MVs erupt hot lava, but watery-mud, methane, CO2, nitrogen, sulfur, and other gases and debris; a mix that could provide nourishment for innumerable organisms (reviewed by Joseph et al. 2020c). Moreover, Parnell and colleagues (2018) have suggested that moist sediments could provide potential habitats for subsurface sulphate reducing organisms, whereas the detection of jarosite supports the possibility of water-atmosphere-biological interactions; i.e. microbially sulphide oxidation (Norlund et al. 2010). On Earth, MV are inhabited by anaerobic methanotrophic archaea, sulfate reducing bacteria, algae and cyanobacteria (Ali et al. 2007; Wrede et al. 2012; Niemann et al. 2006; Remizovschi et al. 2018; Alain et al. 2006) all of which give the erupted mixture a blackish color.
If Martian “mud volcanoes” and geysers are one and the same, is unknown. However, cold water dirty geysers are also believed to contribute to these massive Martian arctic formations (reviewed by Joseph et al 2020c). Cold water geysers are fed by subsurface reservoirs of trapped CO2, geophysically obtained from rock, and the decarbonation of carbonates at depths well below the surface. Subsurface CO2 is also produced via organic decay and aerobic organisms that release CO2 as a waste product (Canfield 2014). As CO2 accumulates, pressures increase, and the geyser erupts water, organic material, and a variety of organisms.
Terrestrial geysers are brimming with life and include sulfur reducers, methanogens and over 100 different microbial phyla representing diverse bacteria and archae (Anantharaman et al. 2016; Probst et al. 2018). If the geysers of Mars are like those of Earth and populated by a wide array of microorganism, then upon erupting they would spray the surface with a variety of organisms, and provide water to those already upon the surface which upon becoming pigmented would turn the surface black.
Radiation: Moulds, Lichens, Fungi, Methanogens, Sulfur Reducing Bacteria
In this and a previous report (Joseph et al. 2020c) we hypothesized and presented evidence of what may be arctic fungus, mould, algae, lichens (a fungal algae composite organism), methanogens and sulfur reducing organisms; all of which may be primarily responsible for the creation of these giant black formation on Mars. We have theorized that methanogens and sulfur reducing species may be ejected onto the surface by geysers and mud volcanoes, and have raised the possibility they all these putative arctic organisms might form spores, migrate beneath the surface or become dormant or die when water evaporates and freezes.
We have also argued that subsurface organisms might die following prolonged exposure to gamma and UV radiation. By contrast, it can be predicted that surface dwelling organisms would have adapted to the radiation intense environment of Mars which has been estimated to equal "0.67 millisieverts per day" at ground level (Hassler et al. 2013). This is significantly and profoundly below the radiation tolerance levels of a variety of prokaryotes (Moseley & Mattingly 1971; Ito et al. 1983) and 4 simple eukaryotes, including fungi and slime moulds which can withstand radiation doses up to 1.7×10 Gy (Deering et al. 1972; Saleh et al. 1988; Terzi, 1961, 1965).
Moulds, fungi, algae, and lichens ((Horneck et al. 2002; McLean & McLean 2010; Nicholson et al. 2000; Novikova et al 2016; Onofri et al. 2012, 2018; Sato et al. 2011; Tugay et al. 2006; Sancho et al. 2007; Raggio et al. 2011; Takahashi, et al. 1972), also survive direct exposure to the radiation intense frigid environment of space with no observable negative effects on viability or spore production. These diverse terrestrial species are well adapted to surviving on Mars. Likewise, it can be predicted that those species that evolved and adapted to the harsh Martian environment would have little difficulty surviving.
Moulds, fungi, lichens, algae, and other species develop adaptive features when exposed to high levels of radiation (Hashimoto & Yanagisawa 1970; Mulleavy & Evans 1982; Rakoczy 1998; Terzi, 1961, 1965; Tugay et al. 2006; Zhdanova et al. 1991, 2004) --a property described as "radiation hormesis" "radiostimulation," and "adiotropism" (Adet 1993; Alshits et al 1981; Levin 2003; Zhuravskaya et al 1995). These adaptations include rapid tissue and cellular regeneration and growth (Basset 1993; Becker 1984; Becker & Sparado 1972; Occhipinti et al. 2014; Levin 2003; Maffei 2014; Moment, 1949) and enhanced spoor production (Hashimoto & Yanagisawa 1970; Rakoczy 1998; Tugay et al. 2006; Zhdanova et al. 1991, 2004).
Putative Martian organisms if provided moisture, would be predicted to grow rapidly in response to the radiation intense Martian environment--and evidence supportive of that hypothesis has been provided in this report. Moreover, lichens, fungi, algae and numerous species of microbe thrive in and are attracted to highly radioactive environments (Becket et al. 2008; Dadachova et al. 2007; Mulleavy & Evans1982; Tugay et al. 2006; Wember & Zhdanova 2001). Fungi (Wember & Zhdanova 2001; Zhdanova et al. 2004) will seek out and grow towards sources of radiation which serve as an energy source for metabolism (Dighton et al. 2008; Tugay et al. 2006). In fact, fungi are invigorated, flourish, and grow rapidly within the International Space Station as a consequence of the heightened radiation levels (Novikova et al. 2016; Novikova 2009; Vesper et al. (2008). Moulds and fungi also flourish on the outskirts and along the walls of the damaged and highly radioactive Chernobyl nuclear power plant (Dighton et al. 2008; Zhdanova et al. 2004). Even if their DNA is damaged by radiation they can easily repair these genes due to a redundancy of genes with repair functions (White et al. 1999). The high levels of radiation, on Mars, may therefore provide an ideal environment for fungi.
CONCLUSIONS
It is well established that a variety of terrestrial organisms survive Mars-like conditions. Given the likelihood Earth has been seeding Mars with life and life has been repeatedly transferred between worlds (Beech et al. 2018; Joseph et al. 2019; 2020c; Schulze-Makuch et al. 2005), it would be surprising if there was no life on Mars. However, in contrast to terrestrial organisms, Martian fungi, lichens, moulds, algae and other putative life-forms, would have evolved on and already be adapted to the low temperatures, intermittent availability of water, low amounts of free oxygen, and high levels of radiation that characterize the harsh Martian environment. Almost all scientists who have searched for current or past life on Mars, have reported positive findings. What would be surprising is if there was no life on Mars.
Positive findings include reports of biological activity and specimens that closely resemble fossilized domical stromatolites, bacterial mats, algae, tube worms, and metazoans; bacterial residue in Martian meteorites; and a host of what may be living fungi, algae, lichens, mould. We have now presented sequential photographs of specimens that emerge from the soil, grow, change shape multiply or move to different locations and disappear; donut shapes resembling the lichen genus Ochrolechia and “milkcap mushrooms” and “donut” shaped Russulas; networks of white specimens resembling plasmodium, bulbous fruiting sporangia and interconnected clumps and sphericals--all of which may have become rigid, encrusted, calcified upon exposure to the surface; as well as specimens resembling puffballs with stalks or shedding what appears to be crustose and which are surrounded by white powder-chunky-leprose-spore-like material that also consists of what appears to be embryonic multi-open-tubular and stemmed mushroom-like formations; and statistical evidence that specimens in the arctic are growing in parallel, and that equatorial spherical specimens resembling fungal puffballs grow out of the ground, expanded in size and diameter, and grew closer together.
Similarities in morphology are not proof of life. It is possible that all the specimens presented here are abiotic. We cannot completely rule out minerals, weathering, and unknown geological forces that are unique to Mars and unknown and alien to Earth. However, growth, movement, alterations in location and shape, constitute behavior, and coupled with life-like morphology, strongly support the hypothesis there is life on Mars.